Research suggests that not only are termites contributing towards climate change, but the changing climate is impacting the the viability of invasive species in new territories.
Latest climate change and termite research
Termite researchers across the world are modelling the potential impacts of climate change on the distribution of termite species. They are also looking at the impact on termite behaviour, in particular the number of days they are likely to remain active (foraging) each year. Researchers have modelled the potential distribution of 15 of the world’s worst invasive pests, under a variety of scenarios. Coptotermes formosanus was the representative termite in this infamous group of insects. Although C. formosanus has the smallest current distribution of the 15 pests evaluated, it had one of the biggest potential shifts in range, no matter the climate scenario. The changing climate would create a highly suitable habitat for C. formosanus over a significantly larger range, with its potential distribution putting much of Asia, Europe, Australia and increasing areas of the US under potential threat.1
The changing climate creates suitable habitats in countries that lack endemic termites – countries with no knowledge of the risk they pose, with pest and construction industries that are simply not set up to deal with the termite threat, should they get introduced and become established. The United Kingdom is an example.
There have only been occasional termite incursions in the UK, and only in the south of England (Devon). Reticulitermes grassei was found on a private property in 1994. The infestation was limited to a few properties, but was only declared eradicated in 2021, after ten years with no reported termite activity. With the government spending a significant amount of time and money on this small eradication project, which nevertheless had the important end goal of preventing termites becoming established, the government is very concerned about the
potential impact should termites become established in the UK. With UK winters typically mild and temperatures warming, recent modelling has indicated that R. flavipes, which is seen as the most likely invasive termite to establish, could survive over much of the UK, including northern England. Its potential distribution may however be impacted by other factors such as soil type.2
An important consideration for pest managers is that even small changes in temperature and rainfall can have a significant impact on termite foraging behaviour, in particular the amount of wood they eat and the duration of the ‘termite season’. The number of days that termites
are active is particularly important for those regions that experience a cold winter, when termite activity can slow or cease completely. Researchers in South Korea inspected 104 wooden architectural buildings across 94 counties in South Korea and found approximately 80%
of them to have termite damage.3 This included some 17 sites where the mean minimum temperature in January was below -4ºC, previously considered an indicative cut off point for suitable habitat for the common termite pest Reticulitermes speratus.

Previous studies had calculated that a 10°C increase in the average annual temperature could lead to an approximately 6.8-fold increase in wood decomposition. The annual mean temperature in South Korea has increased by 1.5°C over the past 100 years, with the average winter minimum having increased by 2.1°C. This has been estimated to have increased the termite activity period by approximately 16 days.
With termite distribution significantly wider than previously thought, and with the period of activity continuing to increase, the authors flagged the need to revisit preventative termite control strategies in South Korea.
References
1 Cao, R. and Feng, J. (2024) ‘Future Climate Change and Anthropogenic Disturbance Promote the Invasions of the World’s Worst Invasive Insect Pests’, Insects, 15(4). https://doi.org/10.3390/insects15040280
2 Brimblecombe, P. (2024) ‘Predicting the changing insect threat in the UK heritage environment’, Journal of the Institute of Conservation, 47(2), pp. 133–148. https://doi.org/10.1080/19455224.2024.2347205
3 Kim, S. and Kim, J. (2024) ‘Estimation of the Damage Risk Range and Activity Period of Termites (Reticulitermes speratus) in Korean Wooden Architectural Heritage Building Sites’, Forests, 15(4). https://doi.org/10.3390/f15040602.
Latest termite dispersal research
Invasive termite species are being discovered in many new locations across the world both by termite researchers and citizen science programs. A warming climate aids in the dispersal of termite species across a wider area but it seems termites are also establishing in cooler locations, finding conditions suitable in sheltered urban areas.
Coptotermes gestroi found in Egypt
Coptotermes gestroi is one of the world’s worst invasive termites and its presence has now been confirmed in Egypt. It was originally misidentified back in 2010 as Reticulitermes lucifugus, before being identified as Coptotermes spp. in 2023, and named C. alexandrines. However, it has now been confirmed that this is indeed C. gestroi.1
Alexandria is the biggest seaport in the Mediterranean and its arrival in Egypt would have been through imported infested goods or more likely from a mating flight aboard an infested boat. This is the most northerly confirmed location of C. gestroi, some 4,000 km away from the nearest confirmed location in northern India. Although the local infestation was extensive, the researchers did not report on the presence of reproductives or other colonies in the area. However, they did note that Alexandria has a climate that could host Coptotermes establishment.

Termites continue to spread in Europe
With the warming temperature, the risk of increasing termite threat in Europe is significant. Reticulitermes grassei is a native termite established in Europe, particularly in Portugal, Spain and France, but also with spot infestations in Switzerland and the UK. Researchers have updated the distribution range of this species using DNA analysis and have identified further locations in Portugal, but also its presence in Morocco and the islands of Madeira and the Azores.2 Their potential impact in urban areas is well understood, although local pest companies currently have little experience with termite management. The researchers indicated that the greater concern would be the impact of invasive termites on island ecosystems.
Invasive termites are also being introduced into cooler northern European countries. As a recent study in Belgium demonstrates, even in countries where cold winters would suggest that certain termite species cannot survive, termites can adapt to the human environment to establish in warmer conditions inside buildings.3 Researchers have confirmed the presence of three invasive species in Belgium: Cryptotermes brevis, Reticulitermes flavipes and R. banyulensis (not considered a pest termite).
The infestation of the West Indian drywood termite, C. brevis, was located in an apartment block in Brussels. It was well established and nuptial flights had been observed for two consecutive years. Although the researchers note that no nuptial flights were observed outside of the building, they note that clearly the microclimate conditions for drywood termites inside buildings allows drywood termite infestations to develop. Whilst less concerned about R. banyulensis the authors were more concerned about the many large nests of R. flavipes found at a zoo in Brugelette. R. flavipes is known to be far more tolerant of cooler temperatures and could become established in Belgium.
The observation of C. brevis has implications for many countries. In Australia for instance, this would mean that drywood termite infestations could potentially establish country-wide in the capital cities and not just beconfined to the warmer areas of the east coast (largely southeast Queensland).
Coptotermes gestroi – the main pest termite in Indonesia?
Coptotermes curvignathus has historically been considered the main pest termite in Indonesia and is the only species specified for inclusion in product registration efficacy testing. However, the distribution and relative abundance of C. curvignathus has long been questioned as C. gestroi is widely distributed in Southeast Asia and is commonly reported in urban areas in Indonesia. A recent paper highlighted that misidentification of C. curvignathus in the field – or rather a lack of identification tools to distinguish between C. curvignathus and C. gestroi – probably provided a false picture of their distribution.4
The researchers concluded that C. curvignathus was the dominant species in forests, whereas C. gestroi was the key threat in urban areas. As a result, they recommended that tests against C. gestroi should be included in registration efficacy testing.
Citizen science discovers drywood termite in South Korea
A homeowner in Seoul posted some photos of termite alates on an internet forum, resulting in an unexpected discovery: the homeowner had an infestation of drywood termites, Cryptotermes domesticus, in a door frame.5 C. domesticus, which is native to Australia and warmer parts of Asia, was not thought to be present in Korea due to the low winter temperatures. Rather than fumigation, the infestation was treated with an insecticide treatment and timber removal, as it appeared to be a localised infestation in the structure. However, since it was a mature colony with alates, ongoing monitoring is in place in Seoul to determine whether this was an isolated colony or C. domesticus is more widely established.
Similarly, in Changwon in southern South Korea, a citizen found alates that were identified as Incisitermes minor, the western drywood termite – an important structural pest native to the US.6 Researchers found that multiple structures around the discovery site had infestations, leading them to conclude that this species of drywood termite could also handle the cooler winter temperatures in South Korea. As it appeared to be well established, the researchers recommended a management plan to prevent further spread.
References
1 Barakat, H. et al. (2024) ‘First record of the invasive Asian subterranean termite, Coptotermes gestroi, from Egypt’, Bulletin of Insectology, 77(1).
2 Hernandez-Teixidor, D. et al. (2024) ‘Molecular identification of the invasive subterranean termite Reticulitermes grassei (Blattodea:Rhinotermitidae) outside its known distribution: introduction routes and implications for pest management strategies’, Journal of Economic
Entomology, 117(3), pp. 1077–1085. https://doi.org/10.1093/jee/toae071
3 Vanderheyden, A. et al. (2024) ‘First record of three alien termite species in Belgium’, Bioinvasions Records, 13(2), pp. 335–344.https://doi.org/10.3391/bir.2024.13.2.04
4 Wikantyoso, B. et al. (2024) ‘The morphological diagnosis of 2 economically important subterranean termites in Western Indonesia,Coptotermes curvignathus and Coptotermes gestroi (Blattodea, Rhinotermitidae)’, Journal of Economic Entomology, 117(5), pp. 2019–2029.https://doi.org/10.1093/jee/toae147
5 Lee, S.-B. et al. (2024) ‘A post in an internet forum led to a discovery of an invasive drywood termite in Korea, Cryptotermes domesticus(Haviland) (Blattodea:Kalotermitidae)’, Journal of Integrated Pest Management, 15(1). https://doi.org/10.1093/jipm/pmae026
6 Lee, S.-B., Jeong, S., et al. (2024) ‘Well-established populations of the western drywood termite, Incisitermes minor (Blattodea:
Kalotermitidae), in Korea’, Journal of Asia-Pacific Entomology, 27(2). https://doi.org/10.1016/j.aspen.2024.102264.
Are termites contributing to climate change?
Termites produce methane as a result of the activity of one of their symbiotic gut microbes, Methanogens archaea. Termite nests are therefore methane hotspots. With methane a potent greenhouse gas, the question is, how much are termites contributing to climate change?
The latest calculations put termite methane emissions at 14.8 Tg methane per year (1 teragram = 1 billion kg). This is about 2% of the total global emissions (natural and human generated) and 4% of the natural emissions.1 Or put another way, the total termite methane emission is more than the human-generated methane emissions released from most countries (on an individual basis), except for the six largest countries.
Of course, termites have been around for millions of years, but when individual countries assess their individual methane emissions, especially for countries with a significant termite biomass, the contribution of termite-related methane may be significant. With countries aiming to reduce methane emissions by 30% by 2030, it is important to know the contribution from termites on a country basis. It is also necessary to acknowledge that as a result of climate change, termite activity – and therefore termite-related methane emissions – will increase. The best case scenario estimated by this latest research is that termite methane emissions will increase by 0.5 Tg/ year, and in the worse-case scenario by 5.9 Tg/year.
References
1 Ito, A (2023). Global termite methane emissions have been affected by climate and land-use changes. Scientific Reports: 13, Article 17195.
Termite dispersal
West Indian drywood termite found in the Sahara
The West Indian Drywood termite, Cryptotermes brevis, is perhaps the world’s most invasive termite, being easily transported within small wooden items. Although it received its name after being discovered in Jamaica in 1845, its natural range is limited to the coastal desert
areas of Peru and Chile. Researchers in Morocco have recently found Cryptotermes brevis in an agricultural area on the western edge of the Sahara Desert (a similar climate to their home range).1 This is the first report of this invasive termite being found outdoors in the ‘Old World’ i.e. within Africa, Asia or Europe.

Reticulitermes Are on The March Too!
Although Coptotermes species and the West Indian drywood termite get most of the attention when it comes to invasive termites, other species are also readily transported by humans. The US has suffered invasions from Coptotermes formosanus, Coptotermes gestroi
and Cryptotermes brevis, but has in turn ‘exported’ Reticulitermes flavipes. Known to be established in Europe, Chile and Uruguay, researchers have now recorded its presence in Buenos Aires, Argentina.2 DNA analysis indicates that it was a separate incursion from the US to those in Chile and Uruguay, and that with the colony producing alates, it is likely to become established.
References
1 Najjari, A. et al. (2023). First outdoor records in the Old World of the invasive drywood termite, Cryptotermes brevis (Walker, 1853) (Kalotermitidae). Journal of Applied Entomology, Short Communication. https://doi.org/10.1111/jen.13156
2 Carrijo, T.F et al. (2023). First record of the major termite pest species, Reticulitermes flavipes (Isoptera: Rhinotermitidae), in Argentina. Journal of Economic Entomology, 116(3): 1033- 1037. https://doi.org/10.1093/jee/toad076
Expanding termite range
Climate change will bring increased average temperatures and higher rainfall globally, although there will be areas of reduced rainfall. Nevertheless, with termites essentially a tropical and semi-tropical animal, it means that the areas suitable for termite colonisation will increase as we move through the century. Even a 1°C increase in average temperature will result in a significant expansion of the areas suitable for termites. Previous modelling on potential Coptotermes distribution has been supported with a recent study on another group of subterranean termites, the Australian harvester termites (Drepanotermes).1 Although not a pest species, they are of interest biologically as they are mainly found in the semi-arid and arid regions of Australia.
The study found that suitable habitat for most species expanded substantially under various climate change scenarios, with only one species having a predicted habitat contraction. However, the researchers made the key point (which applies to all potential termite expansion) that the rate at which climate change is occurring is quicker than the termite dispersal rate, so the range expansion of termite species will likely lag behind the expansion of suitable habitat.
Increased range of food discovery and consumption
One of the other critical impacts of changing climate on termite behaviour is the impact it will have on termite foraging patterns. With increasing temperature termites eat more, so damage can occur more quickly. By replicating an experiment at 133 sites globally, covering a range of temperature and moisture gradients, researchers have attempted to model how termite food discovery and consumption varies with temperature and moisture levels.2 (The research also investigated the effect of decay.)
According to their model, termite discovery of food sources increased with increasing temperature levels (unsurprisingly). However, discovery in relation to moisture levels was more complicated. At higher temperatures, discovery increased with decreasing moisture (rainfall) but at lower temperatures discovery increased with increasing moisture.
With regards to consumption of wood, researchers have found that wood consumption was highly sensitive to temperature, increasing >6.8 times per 10°C in temperature. This figure called the Q10 temperature coefficient is normally around 2 for microbial decay and other biological systems, so a figure of 6.8 is very high, meaning changing temperatures will have a big impact on termite consumption.
The researchers acknowledge that the rate of consumption is impacted by the species that persist in the different climatic areas and the sheer termite numbers, rather than being solely due to the increased rate of consumption by individual termites. Nevertheless, with the likely expansion of geographic range of many termite species, the researchers concluded that the potential for increased termite damage was significant.
In addition to increased consumption rates, especially in the more temperate areas, the ‘termite season’ will extend longer. Importantly, unlike the expansion of termite geographic range, there is no time lag in this change of behaviour; if the temperature changes, the termite foraging behaviour changes, so this is very much something termite professionals should be aware of.
Termite distribution and dispersal
Mapping termite distribution
Detailed information on the termite species present in each country globally is somewhat limited. This is partly due to the cryptic nature of termites and also due to there being an insufficient number of experts who can correctly identify the termite species present. Knowing the termite species present in each country and their distribution within the country is important in identifying potential incursions of invasive termites, and also in understanding how changing distributions within countries could impact control strategies in the future.
Taiwan has carried out a global first: a nationwide termite pest survey managed as a citizen science project.3 Using a Facebook group to help promote the program, samples were sent in by citizens for identification and termite swarming events were also recorded. Over a five-year period (2015-2020) over 3000 samples were collected. Of these, some 1499 were collected from buildings, identifying five structural pests. Of particular interest, the data demonstrated the northward expansion of the invasive termite Coptotermes gestroi and also the first intercepted alate of a Schedorhinotermes species, a potential new invasive pest from Southeast Asia.
Detecting and measuring incursions
It’s important for border security and customs to remain vigilant for potential new termite incursions, as they are certainly happening in many countries around the world. Recent investigations on new infestations of the Formosan termite, Coptotermes formosanus, in southern California, investigated colonies from three new infestation sites compared to two existing ones. Using genetic analysis, the data clearly demonstrated that these five colonies alone had occurred through three separate incursions.4
Can invasive termites interbreed with native species?
Apart from the obvious concern regarding the arrival and establishment of an invasive species, there is also the very real possibility of the invasive termite species interbreeding with native species. In Florida two invasive species are established: Coptotermes formosanus and C. gestroi. Although hybrid colonies have yet to be detected in the field, understanding the potential for such a hybrid is of significant interest and concern. Often hybridisation in social insects leads to ‘hybrid vigour’, where some aspect of their biology provides a competitive advantage for the hybrid over their parents. But is that the case for these Coptotermes hybrids?
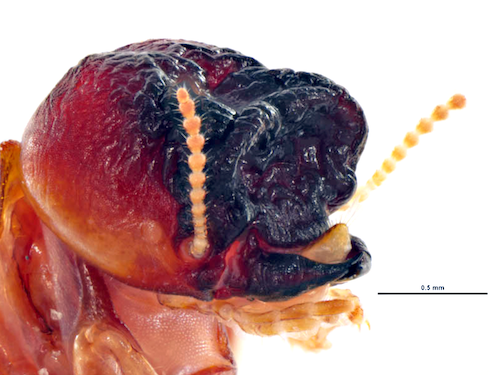
By creating hybrid colonies in the laboratory (main picture, above) researchers in Florida assessed the long-term development over four years.5 The researchers did not find any suggestion of superior hybrid vigour in terms of colony growth when compared to C. formosanus or C. gestroi, but there were no negative effects either, with respect to colony numbers and wood consumption rates. Interestingly, the researchers also discovered that the hybrid colonies produced multiple secondary reproductives, despite the primary reproductive being present – something that rarely occurs in the pure-bred parent species. This may provide the hybrids with an advantage.
Although such hybrids have yet to be detected in the field, this research shows it is definitely possible, if not likely. Furthermore, the hybrid colonies may also have some competitive advantage over their parents. However, it is yet to be determined whether the reproductives from these hybrid colonies are actually fertile, which will certainly affect the assessment of the potential impact of Coptotermes hybrids in Florida.
References
1 Heimburger, Bastian & Soto Maurer, Santiago & Schardt, Leonie & Scheu, S. & Hartke, Tamara. (2022). Historical and future climate change fosters expansion of Australian harvester termites, Drepanotermes. Evolution. 76. 10.1111/evo.14573.
2 Zanne, Amy & Flores-Moreno, Habacuc & Powell, Jeff & Cornwell, William & Dalling, James & Austin, Amy & Classen, Aimee & Eggleton, Paul & Okada, Keiichi & Parr, Catherine & Adair, Carol & Adu-Bredu, Stephen & Alam, Azharul & Alvarez Garzón, Carolina & Apgaua, Deborah & Aragón, Roxana & Ardon, Marcelo & Arndt, Stefan & Ashton, Louise & Zalamea, Paul-Camilo. (2022). Termite sensitivity to temperature affects global wood decay rates. Science. 377. 1440-1444. 10.1126/science.abo3856.
3 Huang, Shih-Ying & Chiu, Chun-I & Tsai, Yi-Ying & Li, Wei-Jie & Wu, Chiachien & Li, Hou-Feng. (2022). Nationwide Termite Pest Survey Conducted in Taiwan as a Citizen Science Project. Journal of Economic Entomology. 115. 10.1093/jee/toac122.
4 Tseng, Shu-Ping & Taravati, Siavash & Choe, Dong-Hwan & Rust, Michael & Lee, Chow-Yang. (2022). Genetic Evidence for Multiple Invasions of Coptotermes formosanus (Blattodea: Rhinotermitidae) in California. Journal of Economic Entomology. 115. 10.1093/jee/toac104.
5 Patel, Jayshree & Lee, Sang-Bin & Chouvenc, Thomas & Su, Nan-Yao. (2023). Equivalent Colony Growth of Hybrids of Two Invasive Coptotermes Species Can Threaten Urban Areas. Journal of Economic Entomology. 116. 10.1093/jee/toad020.
In discussions regarding the impact of climate change on termite behaviour, there is a general perception that a warming environment and changing rainfall patterns will allow termites to increase their distribution. However, it is more complicated than that. Termite physiology and behaviour, and their relationship with their symbiotic gut microbes has evolved over millions of years; how they may respond to a rapidly changing climate is largely unknown and the subject of increasing research.
The impact of temperature
The potential impact of temperature increases is one obvious influencing factor. It’s also important to realise that changes to the climate are likely to impact different species in different ways. Recent research on the termite fauna in Ghana, Africa, has suggested that savanna termite species have wider thermal tolerance limits than forest termite species, which is not surprising given the wider temperature ranges experienced in the more exposed savanna environment.1

The impact of rainfall
It is also incorrect to assume that the same factors drive species distribution in different parts of the world. In most tropical regions, termite abundance and species diversity are assumed to increase with increasing rainfall, with the highest levels of abundance and diversity found in rainforests. However, it appears in Australia that it is the reverse. Assessing species distribution in Australia across savanna, sclerophyll forest and tropical rainforest, researchers found that the highest species diversity and abundance in termites occurred in the low rainfall, savanna sites.2 Only five species were recorded in the rainforests, all wood-feeding species. The researchers concluded that in Australia, other environmental or habitat-related factors shape species distribution.

The impact on gut microbes
In considering the impact of changing climate on termites, much of the focus has been on the potential effect of increasing temperature on the distribution of species and their behaviour, but very little on the impact of their symbiotic gut bacteria, which are critical for their survival. It is hypothesised that since these gut bacteria have evolved along with the termites and therefore have become very specialised, they will also be less adaptable to changes in temperature.
Looking at the impact of temperature on the gut microbial community of Reticulitermes flavipes, researchers measured changes in the makeup of the gut microbe community over a four-week period at three different temperatures: low (15°C), medium (27°C) and high (35°C).3 Whilst the gut microbes in the medium temperature group were similar to field populations and not greatly different to the gut makeup of the low temperature group, the gut microbes present in the high temperature group had changed significantly. Whilst the researchers could not draw any firm conclusions as to the impact of these changes, termite survival in the high temperature group was significantly lower than in the lower temperature groups. The researchers suggested that future work should focus on the potential impact of changes in gut microbe communities on the various termite metabolic processes and behaviours.
Termite species distribution and dispersal
There is an increasing focus on mapping out the global distribution of termite species. Not only is this important in any assessment of the impact of climate change, but it is also important in understanding the risks in further human-mediated transport of invasive termite species, which can have a significant economic and environmental impact.
Mapping species distributions
Whilst many of the pest termite species are well known, it is important to acknowledge that our general understanding of the termite species present in many countries is largely unknown. Using Thailand as an example, researchers have reviewed termite research papers over the last 56 years to compile a list of confirmed termite species.4 Collectively the papers covered 44 defined locations across Thailand. The papers recorded 75 termite species, with the predominant species across all regions being Globitermes sulphureus, Macrotermes gilvus, Microcerotermes crassus, and Microtermes obesi. However, the papers also recorded some 83 unknown species of termites! Mapping of existing termite species is of course important to determine whether an incursion has occurred. An example is the recent discovery of the drywood termite Glyptotermes nakajimai in the Korean Peninsula. Is this a recent incursion, and is it human mediated? 5
Termite incursions and dispersal
Human-mediated movement of goods remains the key mode of travel for invasive pests. This is true not only for drywood termites, which can exist as nests within small pieces of timber, but for subterranean termites too.
Coptotermes gestroi is found in many Asian countries and has also been confirmed in the US and South America. Although officially recorded as present in Northern India, there is some doubt as to whether it is actually present in India.6 However, a number of dead alates were recently identified in a packing box that had arrived from Harrisonburg, Virginia, USA. Although this interception is not expected to have given rise to a successful colony founding (as the alates were all dead), it does indicate that such intrusion pathways exist.
Whilst there will be significant genetic diversity of a species within its native range, the founding colony in an invasion event will have a specific genetic profile. This allows the use of genetic analysis to map the spread of invasive species. Researchers have used this technique to map the spread of Reticulitermes flavipes, by looking at the genetic makeup from 23 native populations in the US and six introduced populations in other countries.
Even within a native range, it is normal for populations to be somewhat isolated geographically and therefore develop a unique genetic profile. However, the researchers established that even within the US, there was quite clear evidence of movement between discrete populations — highly different genetic individuals were found within the same locality and highly similar genetic individuals were found in locations separated by thousands of kilometres.7 This could only be explained by extensive, human-mediated transfer. Whilst this made it more difficult to pinpoint the source of incursions in other countries, the researchers confirmed that the US was the source of incursions in Canada, Chile and France. They further established that the incursion in France became a bridgehead for further introductions into Canada and Chile.
The US is not the only source of termite incursions. Nasutitermes corniger was first detected at Dania Beach in Florida in 2001, probably arriving from South America or the Caribbean.8 However, control attempts did not start until two years later. In 2016 a second population was detected 21 km away and genetic analysis confirmed it to be derived from the original colony in Dania beach, with human transport from one location to the other the likely route of dispersal. As an invasive termite to the US, authorities are focused on eradicating the incursion before it has the chance to establish.

Human-mediated transport of termites is not only an issue between countries but also within countries. Coptotermes formosanus is an invasive termite to the US, with its origin likely in Asia, possibly via Hawaii. It is currently established in the south-eastern states of the US, as far west as Texas. Obviously, it would be ideal to contain its spread as much as possible. However, a recent Coptotermes formosanus infestation was detected in Riverside County, California.9 Whereas two previous incursions in California were determined to have come from within the US, genetic analysis of this new incursion determined that the termites had come from Asia.
Not only subterranean and drywood termites…
Much of the focus on human-mediated termite dispersal has focused on subterranean and drywood termites as their association with wood makes them a more likely candidate for hitchhiking. However, a recent discovery in Japan suggests that soil-nesting, soil-feeding species can also be accidentally transported.10 Pericapritermes nitobei has been recorded on Okinawa Island, despite not being detected during previous extensive termite surveys on the island. The researchers concluded that this was likely a human-mediated incursion. Due to its similarity to the soil-nesting, fungus-growing termite Odontotermes formosanus, which is a pest of forestry and agriculture, the researchers suggested that other non-wood-feeding, non-wood-nesting termites could also be transported by human activity.
Invasive termites can have wide-ranging impacts
Whilst the main concern regarding invasive termites is the potential damage they can do to human structures, termites can also damage agricultural and forestry crops. As an invasive organism, they have the potential to upset the ecological balance in the environments they invade. Evans (2021) notes that about 25% of invasive termites have spread from urban into periurban, forested habitats.11 Although the spread may be slow due to long generation times and short dispersal alate flights, they have the potential to cause significant economic and ecological damage, especially Coptotermes species, which can attack and kill living trees, and develop into large colonies.
References
1 Woon, Joel & Atkinson, David & Adu-Bredu, Stephen & Eggleton, Paul & Parr, Catherine. (2022). Termites have wider thermal limits to cope with environmental conditions in savannas. Journal of Animal Ecology. 91. 10.1111/1365-2656.13673.
2 Clement, Rebecca & Flores-Moreno, Habacuc & Cernusak, Lucas & Cheesman, Alexander & Yatsko, Abbey & Allison, Steven & Eggleton, Paul & Zanne, Amy. (2021). Assessing the Australian Termite Diversity Anomaly: How Habitat and Rainfall Affect Termite Assemblages. Frontiers in Ecology and Evolution. 9. 657444. 10.3389/fevo.2021.657444.
3 Arango, Rachel & Schoville, Sean & Currie, Cameron & Carlos-Shanley, Camila. (2021). Experimental Warming Reduces Survival, Cold Tolerance, and Gut Prokaryotic Diversity of the Eastern Subterranean Termite, Reticulitermes flavipes (Kollar). Frontiers in Microbiology. 12. 10.3389/ fmicb.2021.632715.
4 Lertlumnaphakul, Watthanasak & Ngoen-Klan, Ratchadawan & Vongkaluang, Charunee & Chareonviriyaphap, Theeraphap. (2022). A Review of Termite Species and Their Distribution in Thailand. Insects. 13. 186. 10.3390/insects13020186.
5 Shim, Jaeil & Park, Haechul & Ju, Ho-Jong & Song, Jeong-Hun. (2021). First record of the termite family Kalotermitidae (Blattodea: Termitoidae) in Korea. Journal of Asia-Pacific Entomology. 24. 10.1016/j. aspen.2021.08.017.
6 Venkatesan, Thiruvengadam & Kalleshwaraswamy, C. & Gupta, Ankita & R., Ashika. (2021). Intrusion Pathway of Invasive Asian Subterranean Termite, Coptotermes gestroi (Wasmann) from The Neotropics into The Indian Mainland. Current Science. 120. 1778-1781. 10.18520/cs/v120/ i11/1778-1781.
7 Eyer, Pierre-André & Blumenfeld, Alexander & Johnson, Laura & Perdereau, Elfie & Shults, Phillip & Wang, Shichen & Dedeine, Franck & Dupont, Simon & Bagnères, Anne-Geneviève & Vargo, Edward. (2021). Extensive human-mediated jump dispersal within and across the native and introduced ranges of the invasive termite Reticulitermes flavipes. Molecular Ecology. 30. 10.1111/mec.16022.
8 Thorne, Barbara & Alspach, Sue & Tenn, Katherine & Clark, Marah. (2021). Actions Toward Containment, Control, and Eradication of Invasive Conehead Termites (Blattodea: Isoptera: Termitidae). Journal of Integrated Pest Management. 12. 10.1093/jipm/pmab015.
9 Tseng, Shu-Ping & Boone, Jason & Boone, Lowell & King, Natalee & Taravati, Siavash & Choe, Dong-Hwan & Lee, Chow-Yang. (2021). Genetic Analysis of Formosan Subterranean Termite (Blattodea: Rhinotermitidae) Populations in California. Journal of Economic Entomology. 114. 10.1093/ jee/toab077.
10 Nozaki, Tomonari & Hojo, Masaru & Yashiro, Toshihisa & Matsuura, Kenji. (2021). First Record of the Soil-feeding Higher Termites (Isoptera: Termitidae) from Okinawa Island, the Central Ryukyus, Japan. Sociobiology. 68. 6159. 10.13102/sociobiology.v68i2.6159.
11 Evans, Theodore. (2021). Predicting ecological impacts of invasive termites. Current Opinion in Insect Science. 46. 10.1016/j.cois.2021.03.003.
The impact of climate change on pest populations is becoming a hot topic. With significant changes in temperature, particularly minimum temperatures, and changes in humidity already apparent, the potential spread of termites to new areas previously unsuitable for termite survival is a real possibility.
One example is South Korea and the potential establishment of the significant invasive termite Coptotermes formosanus. Researchers have recently modelled the likelihood of the Formosan termite becoming established in South Korea as a result of climate change.1 Currently, there has been no confirmed record of the Formosan termite in South Korea, despite it being present in two neighbouring countries – China and Japan. This is due to the fact that C. formosanus cannot survive in conditions where the average winter temperature is below 4°C, which was the prevailing situation for the vast majority of South Korea, with the exception of Jeju Island. However, with temperatures increasing, this is no longer the case.
Over the last hundred years, the average annual temperature in South Korea has increased 2.09°C. Some southern areas of South Korea now have a climate that would allow Formosan termites to become established. Assuming the temperature continues to increase at a similar rate, by the end of the century, much of the southern part of South Korea would provide conditions that allow the establishment of C. formosanus.
Critical to avoiding the establishment of C. formosanus is to ensure quarantine inspection prevents its introduction in the first place. With the increasing trade from China, Taiwan, Japan and the US, a southern seaport such as Busan is considered a likely entry point.
Switzerland demonstrated that this is not just a theory, when researchers reported the first record of the subterranean termite, Reticulitermes grassei, in 2020.2 This infestation was first detected in 2018 in a home near Lake Zurich (pictured above). The source of the introduction was assumed to have been olive and palm trees planted on the property, as DNA analysis indicated that the termites were similar to populations from southern Spain. After an initial failed treatment using diatomaceous earth, a baiting treatment appeared to have controlled the infestation. However, the fact that two mating flights had been observed means that authorities need to be vigilant regarding further infestations.
Human-assisted pest invasions are a feature of the modern world. For example, researchers in India recently reported that two non-native Coptotermes species – C. testaceus and C. sjostedti – were intercepted at a quarantine station, having been discovered inside incoming timber.3 Undoubtedly, many unsuccessful invasions see the invading pest either being intercepted or failing to establish; yet some are successful and the chances of success can be improved by the ‘bridgehead effect’. This is where an invasive species establishes a population outside its native range, which then becomes the source for further infestations.
Researchers from the US and China have used DNA analysis to establish that a bridgehead was formed in Hawaii, which provided the source for one introduction of Coptotermes formosanus into the southern US in the 1930s.4 Furthermore, they propose that this bridgehead was actually formed in Hawaii from two separate introductions: one from eastern Asia and one from Hong Kong. Along with a second incursion in Florida directly from South-central China in the 1940s, it allowed C. formosanus to become established in mainland US.
It is also important to appreciate that changes in weather patterns not only allow termites to colonise areas that may have previously been uninhabitable, but may also impact their behaviour within their existing range. Changes in temperature, rainfall and seasonality all have the potential to alter foraging behaviour, reproductive cycles and indeed impact species composition in a given area.
References
1 Lee, S.-B., Tong, R.L., Kim, S.-H., Im, I.G., Su, N.-Y., 2020b. Potential pest status of the Formosan subterranean termite, Coptotermes formosanus Shiraki (Blattodea: Isoptera: Rhinotermitidae), in response to climate change in the Korean Peninsula. FLORIDA ENTOMOLOGIST 103, 431–437.
2 Ghesini, S., Mueller, G., Marino, M., 2020. First record of the subterranean termite Reticulitermes grassei in Switzerland. BULLETIN OF INSECTOLOGY 73, 149–151.
3 Nagaraju, D.K., Kalleshwaraswamy, C.M., Iyyanar, D., Singh, M., Jain, R.K., Kasturi, N., Ranjith, M., Mahadevaswamy, H.M., Asokan, R., n.d. First interception of two wood feeding potential invasive Coptotermes termite species in India. INTERNATIONAL JOURNAL OF TROPICAL INSECT SCIENCE. https://doi.org/10.1007/s42690-020-00287-5
4 Blumenfeld, A.J., Eyer, P.-A., Husseneder, C., Mo, J., Johnson, L.N.L., Wang, C., Grace, J.K., Chouvenc, T., Wang, S., Vargo, E.L., 2021. Bridgehead effect and multiple introductions shape the global invasion history of a termite. COMMUNICATIONS BIOLOGY 4. https://doi.org/10.1038/s42003-021-01725-x
Further reading:
More information on termites.

